EL MONSTRUO DEL LOCH NESS. LOS PRIMOS DE NESSIE (15)
 Durante algún tiempo la verdadera identidad de la carcasa del Zuiyo Maru siguió siendo desconocida Sólo se tenían las mediciones de Michihiko Yano. El cuello medía 1,5 metros de largo; la cabeza, 45 centímetros y las aletas un metro. Del extremo de la cabeza al de lo que parecía ser la cola, la longitud alcanzaba 6 metros. Yano observó que las vértebras bien desarrolladas medían 45 cm y tenía 15 cm de espesor.
Durante algún tiempo la verdadera identidad de la carcasa del Zuiyo Maru siguió siendo desconocida Sólo se tenían las mediciones de Michihiko Yano. El cuello medía 1,5 metros de largo; la cabeza, 45 centímetros y las aletas un metro. Del extremo de la cabeza al de lo que parecía ser la cola, la longitud alcanzaba 6 metros. Yano observó que las vértebras bien desarrolladas medían 45 cm y tenía 15 cm de espesor.
Pero luego, a partir del 26 de julio, se comenzaron a analizar minuciosamente las muestras de tejido que se habían guardado. Varios fueron los  investigadores encargados de estos estudios: el bioquímico doctor Shigeru Kilmora de la Universidad de Tokio, el profesor Fujio Yasuda. En el momento en que el artículo de Oceans iba a la prensa, los científicos en el Japón ya estaban estudiando el caso del Zuiyo-Maru más de cerca. Las copias de las fotografías del canal habían llegado a los
investigadores encargados de estos estudios: el bioquímico doctor Shigeru Kilmora de la Universidad de Tokio, el profesor Fujio Yasuda. En el momento en que el artículo de Oceans iba a la prensa, los científicos en el Japón ya estaban estudiando el caso del Zuiyo-Maru más de cerca. Las copias de las fotografías del canal habían llegado a los  científicos de la Universidad de Pesca de Tokio, incluido su presidente el doctor Tadayoshi Sasaki, quien propuso una reunión de científicos para estudiar los datos disponibles. Las reuniones iniciales fueron el 1 y el 19 de septiembre de 1977, a la que asistieron más de una docena de científicos, entre ellos especialistas en bioquímica, ictiología, paleontología, anatomía comparada, y otros campos. Los científicos acordaron evitar la difusión de sus conclusiones hasta terminar el estudio.
científicos de la Universidad de Pesca de Tokio, incluido su presidente el doctor Tadayoshi Sasaki, quien propuso una reunión de científicos para estudiar los datos disponibles. Las reuniones iniciales fueron el 1 y el 19 de septiembre de 1977, a la que asistieron más de una docena de científicos, entre ellos especialistas en bioquímica, ictiología, paleontología, anatomía comparada, y otros campos. Los científicos acordaron evitar la difusión de sus conclusiones hasta terminar el estudio.
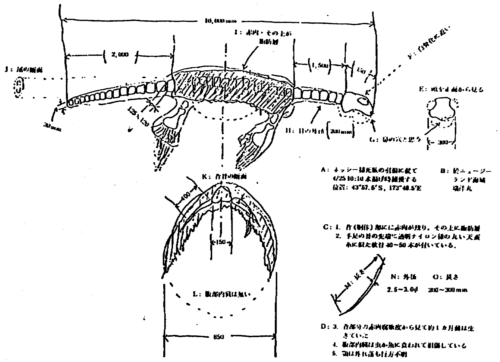 Traducción: A. Captura de una carcasa parecida a Nessie. Capturada el 25 de abril a las 10:00 a.m. a 43 grados 57,5 minutos S, 173 grados 48,5 minutos E (sic). B. En el mar frente a Nueva Zelanda; Zuiyo Maru. C. 1. Restos de músculos rojos en la parte posterior del tronco, superpuestos con capas de grasa. 2. Hay 40-50 piezas de cartílagos transparente, redondos en su sección transversal, parecidos a nylon, alrededor de las puntas y extremidades. D. 3. A juzgar por el estado de putrefacción, el animal pudo haber estado vivo hasta alrededor de un mes antes de su captura. 4. Los órganos internos en el abdomen están dañados, comidos por los gusanos o peces. 5. La mandíbula inferior se ha perdido. E. Vista frontal de la cabeza (300 mm). F. Bien esqueletizado. G. Probablemente fosas nasales (sic). H. Diámetro del hueso vertebral del cuello (200 mm). I. Músculos rojos; capas de grasa en ellos. J. Sección transversal de la cola. K. Sección transversal de la parte posterior del hueso (150 mm). L. No hay órganos internos en el abdomen. M. Longitud. N. Diámetro (¿de las fibras corneas?]. O. Longitud (de las fibras) (200 – 300 mm).
Traducción: A. Captura de una carcasa parecida a Nessie. Capturada el 25 de abril a las 10:00 a.m. a 43 grados 57,5 minutos S, 173 grados 48,5 minutos E (sic). B. En el mar frente a Nueva Zelanda; Zuiyo Maru. C. 1. Restos de músculos rojos en la parte posterior del tronco, superpuestos con capas de grasa. 2. Hay 40-50 piezas de cartílagos transparente, redondos en su sección transversal, parecidos a nylon, alrededor de las puntas y extremidades. D. 3. A juzgar por el estado de putrefacción, el animal pudo haber estado vivo hasta alrededor de un mes antes de su captura. 4. Los órganos internos en el abdomen están dañados, comidos por los gusanos o peces. 5. La mandíbula inferior se ha perdido. E. Vista frontal de la cabeza (300 mm). F. Bien esqueletizado. G. Probablemente fosas nasales (sic). H. Diámetro del hueso vertebral del cuello (200 mm). I. Músculos rojos; capas de grasa en ellos. J. Sección transversal de la cola. K. Sección transversal de la parte posterior del hueso (150 mm). L. No hay órganos internos en el abdomen. M. Longitud. N. Diámetro (¿de las fibras corneas?]. O. Longitud (de las fibras) (200 – 300 mm).
 Yasuda afirmó que había encontrado restos de un aminoácido igual al que se puede encontrar en los restos de los tiburones. Durante su investigación descubrió que las muestras contenían un tipo de proteína muy especial, conocida como elastodin, que sólo se encuentra en los
Yasuda afirmó que había encontrado restos de un aminoácido igual al que se puede encontrar en los restos de los tiburones. Durante su investigación descubrió que las muestras contenían un tipo de proteína muy especial, conocida como elastodin, que sólo se encuentra en los  tiburones.
tiburones.
Como se mencionó, algunos científicos occidentales creían desde el principio que el cadáver en cuestión era probablemente un tiburón, sobre la base de su conocimiento de la decadencia del tiburón peregrino, y otros incidentes similares en el pasado de carcasas de «serpientes de mar». Por otra parte testimonios de algunos marinos apuntaban la confirmación de haber encontrado un escualo de gran tamaño,  ya que según aseguraron el cuerpo únicamente estaba protegido por una especie de esqueleto de cartílago y no huesos como debería haber presentado un plesiosauro.
ya que según aseguraron el cuerpo únicamente estaba protegido por una especie de esqueleto de cartílago y no huesos como debería haber presentado un plesiosauro.
El número de vértebras también correspondía mas con un tiburón que con el esqueleto de un plesiosauro que doblaría  en número el total de vértebras encontradas en el animal hallado. La descripción de las costillas también demostraba el hallazgo de un gran escualo en descomposición.
en número el total de vértebras encontradas en el animal hallado. La descripción de las costillas también demostraba el hallazgo de un gran escualo en descomposición.
 El tiburón peregrino, Cetorhinus maximus, es el segundo pez más grande del mar (sólo superado por el tiburón ballena). Puede crecer a más de 9 metros de longitud, y se han reportado especimenes de más de 12 metros. Sin embargo, este gentil gigante es inofensivo para los seres humanos. Se alimenta
El tiburón peregrino, Cetorhinus maximus, es el segundo pez más grande del mar (sólo superado por el tiburón ballena). Puede crecer a más de 9 metros de longitud, y se han reportado especimenes de más de 12 metros. Sin embargo, este gentil gigante es inofensivo para los seres humanos. Se alimenta  filtrando plancton (en su mayoría minúsculos crustáceos) a través de sus grandes agallas en forma de rastrillo ya que nada lentamente justo debajo de la superficie del agua con la enorme boca abierta. Cuando el tiburón peregrino se desintegra, las mandíbulas y los arcos branquiales a menudo se caen en primer lugar, dejando la aparición de
filtrando plancton (en su mayoría minúsculos crustáceos) a través de sus grandes agallas en forma de rastrillo ya que nada lentamente justo debajo de la superficie del agua con la enorme boca abierta. Cuando el tiburón peregrino se desintegra, las mandíbulas y los arcos branquiales a menudo se caen en primer lugar, dejando la aparición de  un cuello largo y cabeza pequeña. La totalidad o parte de la cola (especialmente en la mitad inferior, que carece de apoyo vertebral) y/o la aleta dorsal también puede desprenderse antes que las mejor unidas aletas pectorales y pélvicas, creando una forma que superficialmente se parece a un plesiosauro. Algunos han llamado a tales restos «pseudoplesiosauro», aunque también se podrían llamar «plesiosharks».
un cuello largo y cabeza pequeña. La totalidad o parte de la cola (especialmente en la mitad inferior, que carece de apoyo vertebral) y/o la aleta dorsal también puede desprenderse antes que las mejor unidas aletas pectorales y pélvicas, creando una forma que superficialmente se parece a un plesiosauro. Algunos han llamado a tales restos «pseudoplesiosauro», aunque también se podrían llamar «plesiosharks».
 La imagen de arriba representa como el cadáver en descomposición de un tiburón peregrino puede dar lugar a algo parecido a un plesiosauro.
La imagen de arriba representa como el cadáver en descomposición de un tiburón peregrino puede dar lugar a algo parecido a un plesiosauro.
A. Tiburón peregrino con la boca cerrada.
B. Tiburón peregrino en el momento de alimentarse.
C. El cuerpo descompuesto del tiburón puede asemejarse a un plesiosauro.
 Como relató el renombrado cryptozoólogo Bernard Heuvelmans, se encontró que más de una docena de canales de supuestas «serpientes de mar» de los últimos años eran definitiva o probablemente cadáveres de tiburón – en la mayoría de los casos de tiburones peregrinos. Estas incluyen, la famosa «Bestia de Stronsa» de las Islas Orkney, Inglaterra (1808), el canal de la Bahía de Raritan New Jersey (1822), la carcasa de la Isla de Henry, Columbia Británica (1934) y el monstruo Querqueville, Francia, también en 1934. Estos fueron seguidos por el canal de Hendaya, en Francia (1951), el canal
Como relató el renombrado cryptozoólogo Bernard Heuvelmans, se encontró que más de una docena de canales de supuestas «serpientes de mar» de los últimos años eran definitiva o probablemente cadáveres de tiburón – en la mayoría de los casos de tiburones peregrinos. Estas incluyen, la famosa «Bestia de Stronsa» de las Islas Orkney, Inglaterra (1808), el canal de la Bahía de Raritan New Jersey (1822), la carcasa de la Isla de Henry, Columbia Británica (1934) y el monstruo Querqueville, Francia, también en 1934. Estos fueron seguidos por el canal de Hendaya, en Francia (1951), el canal  de Nueva Gales del Sur, (1959), y dos casos más en 1961 (Vendee, Francia, y Northumberland, Inglaterra). Se supone que en 1970 el mar arrastró otro «monstruo» a la playa de Scituate,
de Nueva Gales del Sur, (1959), y dos casos más en 1961 (Vendee, Francia, y Northumberland, Inglaterra). Se supone que en 1970 el mar arrastró otro «monstruo» a la playa de Scituate,  Massachusetts. Se dice que esta bestia de 30 pies se parecía a un plesiosauro, pero también resultó ser un tiburón peregrino degradado. En 1996 otra supuesta serpiente de mar había encallado en Block Island, RI. También se ha evaluado como un probable tiburón peregrino, y fue apodado el «Block Ness Monster».
Massachusetts. Se dice que esta bestia de 30 pies se parecía a un plesiosauro, pero también resultó ser un tiburón peregrino degradado. En 1996 otra supuesta serpiente de mar había encallado en Block Island, RI. También se ha evaluado como un probable tiburón peregrino, y fue apodado el «Block Ness Monster».
Curiosamente, los tiburones peregrinos parecen tener una propensión a imitar serpientes marinas en vida, así como muertas. A menudo se alimentan en grupos en o cerca de la superficie (de ahí su nombre), a veces alineados en dos o más filas. Cuando hacen esto, la cola y las aletas dorsales sobresalen de las aguas pueden ser, y, a veces, han sido, confundidos con múltiples «jorobas» y la cabeza de un largo cuerpo de un monstruo de mar.
Los resultados definitivos fueron claros, se trataba de los restos de un tiburón  peregrino de unos 10 metros de longitud y la extraña forma de su cuerpo se debía al hecho de que se hubiese desprendido su mandíbula inferior.
peregrino de unos 10 metros de longitud y la extraña forma de su cuerpo se debía al hecho de que se hubiese desprendido su mandíbula inferior.
El resultado final del estudio fue concluyente, en un comunicado de 9  hojas se detallaron todos los componentes del animal encontrado y se comprobó con una total seguridad que se trataba de un tiburón peregrino en estado de descomposición. Los restos estudiados demostraron ser parte de una estructura cartilaginosa como la que se puede encontrar en el «esqueleto» de los tiburones.
hojas se detallaron todos los componentes del animal encontrado y se comprobó con una total seguridad que se trataba de un tiburón peregrino en estado de descomposición. Los restos estudiados demostraron ser parte de una estructura cartilaginosa como la que se puede encontrar en el «esqueleto» de los tiburones.
En julio de 1978, una  colección de nueve artículos que presentaban las conclusiones del equipo fueron publicados en un informe de la Societe Franco-Japonaise d’Oceanographie. A pesar de algunos desacuerdos sobre temas específicos de las pruebas, y la opinión de algunos trabajadores de que la identificación era todavía incierta, la opinión mayoritaria era que el canal era un tiburón
colección de nueve artículos que presentaban las conclusiones del equipo fueron publicados en un informe de la Societe Franco-Japonaise d’Oceanographie. A pesar de algunos desacuerdos sobre temas específicos de las pruebas, y la opinión de algunos trabajadores de que la identificación era todavía incierta, la opinión mayoritaria era que el canal era un tiburón  descompuesto, y casi todos lo consideraron un tiburón peregrino. Esta conclusión estaba firmemente apoyada en varias líneas de pruebas, incluidos los estudios sobre la apariencia microscópica, la composición química y las propiedades físicas de las muestras de tejidos, así como una serie de consideraciones
descompuesto, y casi todos lo consideraron un tiburón peregrino. Esta conclusión estaba firmemente apoyada en varias líneas de pruebas, incluidos los estudios sobre la apariencia microscópica, la composición química y las propiedades físicas de las muestras de tejidos, así como una serie de consideraciones  anatómicas.
anatómicas.
Si los criptozoólogos siguen pensando que la carcasa del Zuiyo Maru era de un dinosaurio, entonces deberían regresar a sus clases de secundaria porque no existen dinosaurios marinos. Animales como los extintos plesiosauros eran saurios marinos pero no eran dinosaurios al  igual que los pteradones eran saurios voladores pero tampoco eran dinosaurios.
igual que los pteradones eran saurios voladores pero tampoco eran dinosaurios.
Los resultados de los estudios científicos, desafortunadamente, no tuvieron la misma difusión que las fotografías y los reportes sensacionalistas. El mejor resumen que he encontrado es el de Glen J. Kuban[1]:
Pruebas de las muestras de tejido
– Las fibras córneas de la muestra del canal eran estructuras rígidas de tipo aguja cónica que hacia los extremos tenían un color  marrón translúcido. Esos rasgos son característicos de ceratotrichia, la fibra cartilaginosa de las aletas de los tiburones raya. Abe encontró que las fibras del
marrón translúcido. Esos rasgos son característicos de ceratotrichia, la fibra cartilaginosa de las aletas de los tiburones raya. Abe encontró que las fibras del  canal, y las ceratotrichia de la carcasa de un tiburón peregrino «se asemejaba notablemente entre sí».
canal, y las ceratotrichia de la carcasa de un tiburón peregrino «se asemejaba notablemente entre sí».
– El análisis de aminoácidos de las muestras del canal dio resultados que concordaban con la elastoidina de un tiburón peregrino. La elastoidina colagenosa es una proteína que sólo se conocen en los tiburones y las rayas. La concordancia era especialmente impresionante cuando la elastoidina de un tiburón peregrino fue tratada con una solución antiséptica de hipoclorito de sodio (NaClO), al igual que las muestras del Zuiyo-Maru.  La correspondencia es prácticamente idéntica en todos los 20 aminoácidos probados (Tabla 1). Al examinar esta «sorprendente similitud», Kimura, Fujii, y otros señalaron que una prueba estadística denominada «Ãndice Diferencial (DI)» dio el valor extremadamente bajo de .95 indicando una apretada correspondencia. También señalaron que el alto contenido de tirosina (43 y 41 para los residuos de las muestras) es especialmente característico de la
La correspondencia es prácticamente idéntica en todos los 20 aminoácidos probados (Tabla 1). Al examinar esta «sorprendente similitud», Kimura, Fujii, y otros señalaron que una prueba estadística denominada «Ãndice Diferencial (DI)» dio el valor extremadamente bajo de .95 indicando una apretada correspondencia. También señalaron que el alto contenido de tirosina (43 y 41 para los residuos de las muestras) es especialmente característico de la ![]() elastoidina del tiburón en comparación con otros colágenos, que normalmente tienen 5 o menos residuos. Ceratotrichia.
elastoidina del tiburón en comparación con otros colágenos, que normalmente tienen 5 o menos residuos. Ceratotrichia.
| Aminoácido | Carcasa de 1977 | Muestra de elastoidina de un tiburón peregrino |
| 4-Hidroxiprolina | 45 | 45 |
| Ãcido aspártico | 54 | 55 |
| Theonina | 25 | 25 |
| Serina | 39 | 40 |
| Ãcido glutámico | 80 | 80 |
| Prolina | 130 | 125 |
| Glicina | 291 | 290 |
| Alanina | 109 | 110 |
| Cistina (1/2) | 7 | 6 |
| Valina | 25 | 24 |
| Metionina | 10 | 10 |
| Isoleucina | 20 | 20 |
| Leucina | 19 | 19 |
| Tirosina | 43 | 41 |
| Fenilalanina | 12 | 12 |
| hidroxilisina | 5 | 6 |
| Lusina | 25 | 26 |
| Histidina | 11 | 13 |
| Arginina | 51 | 53 |
| (Amida-N) | (57) | (62) |
Cuadro 1. Resultados del análisis de aminoácidos de las fibras córneas de la carcasa de 1977 del Zuiyo Maru y la elastoidina de un tiburón peregrino (residuos/1000 residuos). Composición fue determinada por cromatografía líquida JLC-3BC (JEOL Co Ltd). Ambas muestras habían sido tratadas con NaClO. (Kimura, Fujii, y otros 1978).
– Las fibras córneas de la aleta mostraron una disminución distintiva de aproximadamente 1/3 del tamaño original cuando se calentaron en agua a 63 grados C, y poco a poco se reelongaron con refrigeración. Este único comportamiento hidrotermal es característico de la elastoidina (Kimura, Fujii, y otros de 1978, p 68).
– Micrografías electrónicas de los tejidos mostraron numerosas protofibrillas paralelas, junto con un patrón de bandas que es característico de la elastoidina del tiburón. Las micrografías también revelaron un importante patrón de estriación periódico de 450-500 angstroms, que es más corto que los colágenos típicos, pero que ya e había observado en la elastoidina del tiburón peregrino (Kimura, Fujii, y otros 1978).
– Los `primeros análisis de cromatografía de gases de las fibras córneas dieron resultados coherentes con los tejidos de tiburones (Sasaki 1978).
Kimura, Fujii, y otros (1978) llegaron a la conclusión de que las muestras estudiadas de tejido compuesto indicaban que las fibras córneas eran esencialmente idénticas a la elastoidina del tiburón peregrino tanto en su morfología y composición de aminoácidos. Se comentó, «Si las fibras córneas se retiraron de animales pertenecientes a otras clases, excepto Chondrichthyes (parientes de los tiburones), deberían ser significativamente diferentes… Estos resultados sugieren fuertemente que esta criatura no identificada es un tiburón peregrino o estrechamente relacionado con la especie (Kimura, Fujii, y otros de 1978, p 73).
Anatomía
– El boceto del canal mostraba seis vértebras del cuello, consideradas como «siete o algo así» por Obata y Tomoda (1978), que es razonablemente coherente con las mediciones del largo cuello hechas por Yano (150 cm) y el diámetro de cada vértebra (20 cm). También es coherente con los tiburones. Sin embargo, las 6 a 7 vértebras cervicales no es coherente con los plesiosauros y otros reptiles marinos. Incluso el pliosaurio, también conocido como plesiosauro «de cuello corto», tienen al menos 13 vértebras en el cuello, el plesiosauro de «cuello largo» tiene muchas más. (Obata y Tomoda, 1978, p 46).
– Se informó que la cabeza de la criatura era como la de una tortuga (Obata y Tomoda, 1978, p 48). Esto es coherente con los restos craneales de un tiburón peregrino, que han sido específicamente descritos como parecidos a la cabeza de una tortuga (Omura, Mochizuki, y Kamiya 1978, p 59). En contraste, la cabeza del plesiosauro tenía una forma triangular, que no eran particularmente parecida a la de un tortuga (Hasegawa y Uyeno 1978, p 64).
– Las fotografías y los testigos confirman la presencia de rayos en las aletas, que son poseídos por la mayoría de los peces, incluidos los tiburones. En cambio, los plesiosauros tenían falanges óseas como soportes de las aletas, que no fueron vistos en la carcasa (Obata y Tomoda 1978, p 51). Los huesos de las extremidades mostrados en el dibujo de Yano, evidentemente, se basan en la presunción de que se trata de un plesiosauro pero no en lo observado (Omura y otros de 1978, p 56; Obata y Tomoda 1978, p 49).
– Una de las fotos (Figura 1c), muestra una aparente aleta dorsal, (como se ilustra en la figura 5). La mayoría de los peces tienen aletas dorsales, incluyendo tiburones, pero se cree que los plesiosauros no las tenían.
– Las formas en «V» a lo largo de la columna vertebral (Figura 1c y 5), y cerca de la faja pectoral (Figura 1a) fueron identificadas como myocommata por Omura, Mochizuki, y Kamiya 1978, p 56-57). Los Myocommata se componen principalmente de tejido conectivo entre los myomeres, y se encuentran en los tiburones, pero no en los reptiles.
– Las costillas medían unos 40 cm de largo, que son demasiado cortas para los plesiosauros u otros vertebrados marinos pero no para los tiburones (Hasegawa y Uyeno, p 65). Irónicamente, algunos se han preguntado si las costillas podrían ser demasiado largas para un tiburón, que suele tener costillas muy pequeñas. Pero se trataba de un espécimen excepcionalmente grande, y probablemente era aún mayor antes de la descomposición. Además, no es seguro que Yano identificara o midiera con precisión las costillas, que no aparecen en las fotos. Quizá por error midió los restos de los arcos branquiales, myocommata, o surcos musculares, y supuso que correspondían a las costillas.
– Como se ve en las fotos, las aletas anteriores parecen estar articuladas en ángulo recto con el hombro, en consonancia con los tiburones, pero no con los plesiosauros (Obata y Tomoda 1978, p 46); Hasegawa y Uyeno 1978, p 65). La faja pectoral es visible entre las aletas en la parte delantera Figuras 1a y 1b, y parece que está rota, pero su forma es parecida a la de los tiburones (Compagno 1997; Phelps 1997; Roesch 1997).
– Si el cadáver eran de un plesiosauro, el cuerpo no podría doblarse en la postura que aparece en algunas de las fotografías, ya que los huesos del pecho serían grandes y planos. Asimismo, los huesos ventrales del plesiosauro tendrían que haber permanecido si las aletas anteriores se conservaron, pero no se ven en la carcasa (Hasegawa y Uyeno 1978, p 64).
– En los plesiosauros, todos los huesos de las extremidades se encuentra en la parte ventral (inferior) del cuerpo, por lo que si la criatura fuera un plesiosauro degradado, es probable que los miembros ya hayan sido separados del cuerpo (Hasegawa y Uyeno 1978, P 63).
– En el actual grado de descomposición, un plesiosauro probablemente habría conservado su mandíbula superior y los dientes (Hasegawa y Uyeno 1978, p 63), pero no se informó de dientes en el canal (Obata y Tomoda 1978, p 48). No obstante, se sabe que un tiburón peregrino, ambas mandíbulas se sueltan fácilmente, y aunque se mantiene el maxilar superior, sus extremadamente pequeños dientes podrían haber sido fácilmente pasados por alto.
– El cadáver fue reportado como de una longitud de 10 metros. Los tiburones peregrinos comúnmente crecen más de 9 metros (Dingerkus 1985; Freedman 1985), y se han reportado especimenes de más de 12 pies de largo (Heuvelmans 1968; Herald 1975; Soule 1981; Acero 1985). Algunos autores indican que incluso pueden crecer más de 15 metros (Springer y oro 1989; Perrine 1995; Allen 1996), El tamaño del canal también sería compatible con un pequeño plesiosauro, pero el cuerpo no es proporcional (se explica más adelante).
– A pesar de que algunas mediciones de Yano parecen sorprendentemente redondeadas (por ejemplo, 2000 mm de la cola y 10000 mm de longitud total), si asumimos que son razonablemente precisos, entonces las proporciones del cuerpo (aproximadamente 2:6:2 para la cabeza+cuello:torso:cola) son incompatibles con cualquier fósil de plesiosauro conocido (Obata y Tomoda 1978, p 52). En muchos plesiosauros el cuello es la sección más larga, y en ningún caso el tronco (entre las aletas pectorales y la pelvis) es más largo que la cabeza y el cuello, como lo es en la carcasa. El canal podría haber perdido algo de su longitud a través de la pérdida de la cola (que se mencionan a continuación), pero la relación cuello – tronco aún sería incompatible con los plesiosauros.
– Las proporciones corporales del canal son, en gran medida compatibles con un gran canal de un tiburón peregrino, sobre todo uno que perdió su cola (comparar Figuras 5 y 2). Sería probable la pérdida de la cola, ya que la gran cola tendería a golpear en la estrecha coyuntura durante la decadencia en el agua. Esto explicaría la burda disminución, en lugar de cola, en el esquema de Yano. El rostrum (punta de la nariz) también puede haberse perdido, pero no afecta de forma apreciable a la longitud del cuerpo o las proporciones. Añadir una cola significaría que el tiburón estaba más cerca de 12,5 metros en vida, que sería excepcionalmente grande, pero todavía dentro del rango de aceptación del tamaño general de los tiburones peregrinos. Después de todo, este pobre peregrino pudo haber muerto de vejez.
La combinación de pruebas anatómicas por lo tanto, indican firmemente un tiburón y desechan eficazmente un plesiosauro. Obata y Tomoda (1978, p. 52) concluyen, «no se conocen fósiles de reptiles que se parezcan al animal examinado». Asimismo, Uyeno y Hasegawa (1978, p. 64) escriben, «Desde el punto de vista osteológico, llegamos a la conclusión de que esta criatura no pertenece a los reptiles plesiosauros».
Varias Observaciones
– Procesadores japoneses de aletas de tiburón, que conocen cabalmente los cadáveres de tiburón, identifican el animal en las fotografías de Yano como un tiburón (Abe 1978).
– En septiembre de 1977, el canal de un tiburón peregrino positivamente identificado había encallado en Nemuro, Hokkaido, y mostraba una notable semejanza con el cadáver del Zuiyo-Maru encontrado tan sólo cinco meses antes. Al describir el animal varado en septiembre Omura, Mochizuki, y Kamiya (1978, p 59-60), escribieron, «Las mandíbulas y arcos branquiales han desaparecido, y el cráneo tiene una apariencia como de tortuga… las aletas pectorales y pélvicas estaban dañadas en sus apexes, pero seguían permaneciendo. Los resultados de este experimento realizado por la naturaleza apoyan la opinión de que el canal del Zuiyo-Maru era un tiburón gigante que había perdido sus mandíbulas y arcos branquiales».
Resumiendo sus conclusiones, Hasegawa y Uyeno (1978) establecen, «Sobre la base de las pruebas disponibles, estamos convencidos de que esta criatura de Nueva Zelanda no es el «Nuevo Nessie», que gran parte del mundo está esperando, sino probablemente sólo un cadáver perteneciente a un tiburón de gran tamaño».
Las presuntas incoherencias
A pesar de todas las pruebas que apuntan a un tiburón, en el informe de 1978 y en otros lugares fueron planteadas algunas incoherencias con la supuesta identificación de tiburones, y también deben revisarse.
– El canal reportado olía como un mamífero marino muerto, y carecía de un olor característico del amoníaco de los cadáveres de tiburón (Hasegawa y Uyeno 1978, p 65). Sin embargo, no se sabe si todos los tiburones desprender olor de amoniaco cuando están descomponiéndose, o por cuánto tiempo. Los mismos autores señalaron que la falta de olor a amoníaco podría ser debida a la magnitud de la pérdida y la descomposición de la piel, de modo que el amoníaco del canal fue lavado por el mar (Hasegawa y Uyeno 1978, p 65). Además, se sabe que incluso vivos, los tiburones peregrinos emiten un singular y altamente ofensivo olor propio (Steel 1985; Ellis 1989), que podría haber dominado todo el olor de amoníaco.
– Una sustancia blanca, pegajosa, como grasa cubría la mayor parte del canal (Obata y Tomoda 1978, p 49). Aunque Niermann (1994, p 103) y algunos otros (Hasegawa y Uyeno 1978) consideran este como el más fuerte argumento en contra de la teoría del tiburón, es realmente compatible con ella. Los tiburones peregrinos tienen grandes depósitos de grasa en el músculo blanco y el hígado. Según algunas autoridades aumentan las reservas de grasa durante el verano, para usarla en el invierno (Steel 1985; Sims 1997). El animal en cuestión probablemente falleció a finales de marzo o principios de abril, que es final del verano, en Nueva Zelanda. Además, uno de los trabajadores japoneses (Seta 1978) explica los fenómenos de la formación de adipocere en cadáveres de tiburones y otros animales en descomposición, en virtud del nuevo material graso que se puede generar durante el proceso de decaimiento. Seta indicó que la sustancia blanquecina, de olor pútrido, viscosa en la carcasa es compatible con la formación de adipocere. También, algo del material blanquecino, probablemente consistía de ligamentos y tejido conectivo filamentosos (Omura, Mochizuki, y Kamiya 1978, p 56). Esos tejidos fibrosos en otros canales tiburón peregrino evidentemente han llevado a algunos informes de cadáveres de «monstruos marinos» con melena de pelo blanco (Heuvelmans 1968; Sweeney 1972).
– Las fotografías muestran, la presencia de músculo rojizo bajo el material blanco, que Obata y Tomoda (1978, p49) sugieren es compatible con un tetrápodo (animales de cuatro patas). Sin embargo, la presencia de músculo rojizo también es compatible con un tiburón. Los tiburones al igual que otros peces tienen músculos blancos y rojos (Fowler 1997; Rey 1997; Sims 1997). El blanco predomina, pero los peces que nadan lentamente y de manera constante, como los tiburones peregrinos en general, tienen más músculo rojo que otros tiburones (Tullis 1997). Algo del color rojizo también puede ser debido a los residuos de sangre.
– Las preocupaciones de algunos autores acerca de la «pequeña cabeza» o «cuello largo» (Koster 1977, Yasuda y Taki 1978) se eliminan una vez que se entiende el proceso del decaimiento en los tiburones peregrinos. Omura, Mochizuki, y Kamiya (1978, p. 59), resumen este proceso, «… un pequeño cráneo y un cuello desproporcionadamente largo y delgado, se explican por la pérdida de las mandíbulas y de los arcos branquiales, en el transcurso de la  descomposición del canal».
descomposición del canal».
– Obata y Tomoda (1978, p 48) también sugieren que, a diferencia de los tiburones, en la que los nares (orificios nasales) se encuentran en la superficie inferior del cráneo, el cadáver tenía agujeros que Yano llamó «probables nares» en la parte delantera del  cráneo. Sin embargo, el rostrum o la mayor parte de la estructura anterior pudo haber desaparecido, por lo que los nares podrían haber estado en la cara inferior y también el «frente» de lo que quedaba del cráneo, eliminando cualquier incompatibilidad. O bien, lo qué Yano cree que son los nares podrían haber sido cualquiera de
cráneo. Sin embargo, el rostrum o la mayor parte de la estructura anterior pudo haber desaparecido, por lo que los nares podrían haber estado en la cara inferior y también el «frente» de lo que quedaba del cráneo, eliminando cualquier incompatibilidad. O bien, lo qué Yano cree que son los nares podrían haber sido cualquiera de  varios otros orificios fenestrales que existen en los cráneos de tiburones, u otros nuevos creados durante la descomposición.
varios otros orificios fenestrales que existen en los cráneos de tiburones, u otros nuevos creados durante la descomposición.
– Algunos testigos negaron la presencia de una aleta dorsal (Obata y Tomoda 1978). Sin embargo, incluso si no había una aleta dorsal, podría haber  estado podrida. En segundo lugar, como se mencionó, una foto muestra una aparente aleta dorsal (ver Figuras 1c y 5), que evidentemente, fue pasada por alto por Yano y otros. Omura, Mochizuki, y Kamiya (1978, p 56), «… por un minucioso examen de la fotografía podemos distinguir claramente la
estado podrida. En segundo lugar, como se mencionó, una foto muestra una aparente aleta dorsal (ver Figuras 1c y 5), que evidentemente, fue pasada por alto por Yano y otros. Omura, Mochizuki, y Kamiya (1978, p 56), «… por un minucioso examen de la fotografía podemos distinguir claramente la  base de una aleta dorsal, a pesar de que ha disminuido desde mediados de la línea dorsal». Señalan que ésta un poco dislocada aleta dorsal evidentemente se había superpuesto parcialmente a la aleta pectoral derecha, que puede dar cuenta de la descripción de Yano de estas últimas como teniendo dos tipos de fibras
base de una aleta dorsal, a pesar de que ha disminuido desde mediados de la línea dorsal». Señalan que ésta un poco dislocada aleta dorsal evidentemente se había superpuesto parcialmente a la aleta pectoral derecha, que puede dar cuenta de la descripción de Yano de estas últimas como teniendo dos tipos de fibras  córneas.
córneas.
– Obata y Tomoda (1978, p. 49) sugieren que las «costillas largas y cilíndricas» en el canal no se encuentran en selachians. «Sin embargo, como se explicó anteriormente, no es seguro que Yano identificara o midiera con precisión las costillas. Incluso Si lo hizo, la longitud de la costilla (40 cm), es más compatible con un gran tiburón que con un plesiosauro. Si la criatura fuera un plesiosauro, habría tenido que ser un plesiosauro de cuello corto, cuyas costillas serían al menos el triple de la longitud informada (John Martin 1997).
– Se dijo que la cabeza  era muy dura, mientras que los tiburones no contienen huesos, sólo esqueletos de cartílago. Sin embargo, el cartílago en los cráneos de tiburón puede ser muy duro y denso, y los tiburones peregrinos tienen esqueletos especialmente bien calcificados (Steel 1985). También, cuando envejece un tiburón, su cráneo se vuelve muy duro y denso. El tamaño de la carcasa claramente indica un espécimen viejo.
era muy dura, mientras que los tiburones no contienen huesos, sólo esqueletos de cartílago. Sin embargo, el cartílago en los cráneos de tiburón puede ser muy duro y denso, y los tiburones peregrinos tienen esqueletos especialmente bien calcificados (Steel 1985). También, cuando envejece un tiburón, su cráneo se vuelve muy duro y denso. El tamaño de la carcasa claramente indica un espécimen viejo.
– Algunos de los miembros de la tripulación dijeron que las aletas pelvianas (posteriores) eran similares en tamaño a las aletas pectorales, como en un plesiosauro (Obata y Tomoda 1978, p 49). Sin embargo, esto no se puede confirmar, ya que  no hay mediciones ni se tomaron fotos de las aletas pélvicas. Yano y otros pudieron haber confundido la gran aleta dorsal, colgando y dislocada con las otras aletas (Hasegawa y Uyeno, (1978, p 62). O bien, la combinación de las aletas pélvica trasera y los broches genitales crearon la ilusión de una
no hay mediciones ni se tomaron fotos de las aletas pélvicas. Yano y otros pudieron haber confundido la gran aleta dorsal, colgando y dislocada con las otras aletas (Hasegawa y Uyeno, (1978, p 62). O bien, la combinación de las aletas pélvica trasera y los broches genitales crearon la ilusión de una  gran aleta trasera (Hasegawa y Uyeno 1978, p 63). Esto podría explicar el comentario de Yano de que la parte trasera tenían una inusual apariencia como la de una foca
gran aleta trasera (Hasegawa y Uyeno 1978, p 63). Esto podría explicar el comentario de Yano de que la parte trasera tenían una inusual apariencia como la de una foca  (Koster 1977). También es posible que las aletas pectorales se degradaron un poco más que las aletas pélvicas, reduciendo la disparidad de su tamaño. El mismo Yano reconoció que haciendo su mejor esfuerzo para recordar las aletas frontales eran algo más grandes que las traseras (Koster 1977). Su boceto sugiere otra cosa, pero se sabe que contiene una serie de inexactitudes, como los huesos de las aletas que realmente no se vieron. Tomando nota de esos problemas, Yasuda y Taki (1978) consideraron el dibujo inherentemente poco fiable, y Obata y Tomoda (1978) sugieren que fue influenciado por los prejuicios. En efecto, en el tiempo en que Yano hizo
(Koster 1977). También es posible que las aletas pectorales se degradaron un poco más que las aletas pélvicas, reduciendo la disparidad de su tamaño. El mismo Yano reconoció que haciendo su mejor esfuerzo para recordar las aletas frontales eran algo más grandes que las traseras (Koster 1977). Su boceto sugiere otra cosa, pero se sabe que contiene una serie de inexactitudes, como los huesos de las aletas que realmente no se vieron. Tomando nota de esos problemas, Yasuda y Taki (1978) consideraron el dibujo inherentemente poco fiable, y Obata y Tomoda (1978) sugieren que fue influenciado por los prejuicios. En efecto, en el tiempo en que Yano hizo  su dibujo (dos meses después de la captura) la idea del plesiosauro se había vuelto tan popular y Yano se había convertido en algo más que una celebridad (Koster 1977).
su dibujo (dos meses después de la captura) la idea del plesiosauro se había vuelto tan popular y Yano se había convertido en algo más que una celebridad (Koster 1977).
– Algunos lectores se estarán preguntando si la ubicación del hallazgo es un problema para la identificación del tiburón peregrino (tal como insinuaron Yasuda y Taki 1978). Sin embargo, se sabe que los tiburones peregrinos habitan muchas partes templadas del mundo, incluyendo las aguas que rodean a Nueva Zelanda (Burton y Burton de 1969; Springer y Gold 1989; Francis 1997). Se pensaba que el animal había muerto en una zona al sur del sitio de captura, bien conocido dentro de la gama de tiburones peregrinos (Nasu 1978).
Los monstruos no mueren fácilmente
 Todos los informes de 1978 muestran una fuerte evidencia a favor de la identificación del tiburón, y no hay objeciones sustanciales a la misma. Incluso autores como Obata y Tomoda, quien inicialmente apoyaron la idea del plesiosauro e hicieron hincapié en los posibles problemas para la interpretación del
Todos los informes de 1978 muestran una fuerte evidencia a favor de la identificación del tiburón, y no hay objeciones sustanciales a la misma. Incluso autores como Obata y Tomoda, quien inicialmente apoyaron la idea del plesiosauro e hicieron hincapié en los posibles problemas para la interpretación del  tiburón, reconocieron que la mayoría de las pruebas se referían a un tiburón y excluían al plesiosauro. Manifestaron que, «No se conocen fósiles de reptiles que concuerden con el animal en cuestión» (Obata y
tiburón, reconocieron que la mayoría de las pruebas se referían a un tiburón y excluían al plesiosauro. Manifestaron que, «No se conocen fósiles de reptiles que concuerden con el animal en cuestión» (Obata y  Tomoda 1978). La mayoría de los otros autores de los informe de 1978 indican más claramente que las pruebas indicaban fuertemente un tiburón peregrino o especies estrechamente relacionadas (Abe, 1978; Hasegawa y Uyeno 1978; Omura, Mochizuki, y Kamiya 1978; Kimura et al 1978).
Tomoda 1978). La mayoría de los otros autores de los informe de 1978 indican más claramente que las pruebas indicaban fuertemente un tiburón peregrino o especies estrechamente relacionadas (Abe, 1978; Hasegawa y Uyeno 1978; Omura, Mochizuki, y Kamiya 1978; Kimura et al 1978).
 Lamentablemente, los informes de 1978 recibieron menos atención pública que las historias originales del «monstruo marino». Del mismo modo, varios escritores de historias de monstruos y misterios continúan describiendo el caso como no resuelto, incluyendo Welfare y Fairley (1980), Soule (1981), y Bord y Bord (1989). Sin embargo, algunos buenos resúmenes de la investigación de 1978 fueron proporcionadas por Cohen (1982), Bright (1989), LeBlond (1992), y Ellis (1994) que dejan de
Lamentablemente, los informes de 1978 recibieron menos atención pública que las historias originales del «monstruo marino». Del mismo modo, varios escritores de historias de monstruos y misterios continúan describiendo el caso como no resuelto, incluyendo Welfare y Fairley (1980), Soule (1981), y Bord y Bord (1989). Sin embargo, algunos buenos resúmenes de la investigación de 1978 fueron proporcionadas por Cohen (1982), Bright (1989), LeBlond (1992), y Ellis (1994) que dejan de  lado cualquier esperanza de que la bestia sea un plesiosauro, y explican correctamente que la muestra evidentemente representa uno de los varios canales de tiburón peregrino confundidos como un monstruo de mar.
lado cualquier esperanza de que la bestia sea un plesiosauro, y explican correctamente que la muestra evidentemente representa uno de los varios canales de tiburón peregrino confundidos como un monstruo de mar.
Lamentablemente, muchos creacionistas siguieron  promoviendo la interpretación del plesiosauro mucho después de 1978, incluidos Ian Taylor (1984, 1987, 1989, 1996), Paul Taylor (1984, 1987), Baugh (1987), Peterson (1988), Baker (1988), Dye (1989), Bartz (1990, 1992), Buckna (1993) y Morris (1993, 1997). La mayoría parece ignorar la investigación y los informes de 1978.
promoviendo la interpretación del plesiosauro mucho después de 1978, incluidos Ian Taylor (1984, 1987, 1989, 1996), Paul Taylor (1984, 1987), Baugh (1987), Peterson (1988), Baker (1988), Dye (1989), Bartz (1990, 1992), Buckna (1993) y Morris (1993, 1997). La mayoría parece ignorar la investigación y los informes de 1978.  Algunos de plano llaman a la bestia plesiosauro (Scoggan 1996; Hovind 1996), o «monstruo de mar» (Doolan 1994), o «dinosaurio» (Hovind 1996) (los plesiosauros no son dinosaurios). Aún más sorprendente fueron los comentarios de los creacionistas, que parecen conscientes de los trabajos en las muestras de tejidos de 1978, y, sin embargo,
Algunos de plano llaman a la bestia plesiosauro (Scoggan 1996; Hovind 1996), o «monstruo de mar» (Doolan 1994), o «dinosaurio» (Hovind 1996) (los plesiosauros no son dinosaurios). Aún más sorprendente fueron los comentarios de los creacionistas, que parecen conscientes de los trabajos en las muestras de tejidos de 1978, y, sin embargo,  sugieren que apoyan la identificación del plesiosauro. Entre las más inquietantes son las siguientes declaraciones:
sugieren que apoyan la identificación del plesiosauro. Entre las más inquietantes son las siguientes declaraciones:
«A partir de fotografías, croquis con mediciones cuidadosas, y el análisis de muestras de tejido de la aleta, que tenía todo el aspecto de ser un plesiosauro o dinosaurio  que vive en el mar…» (Ian Taylor 1984, 1978)
que vive en el mar…» (Ian Taylor 1984, 1978)
«Fotografías, exámenes de tejidos, y medidas hechas, demostraban que era probablemente un plesiosauro». (Paul Taylor 1987).
«Las fotografías, exámenes de tejidos, y las mediciones fueron hechas por los científicos japoneses. Sus conclusiones apuntan a un descendiente de plesiosauro» (Baker,  1988).
1988).
Algunos incluso se quejaron de que la prensa estaba reprimiendo la historia del plesiosauro (Bartz 1992; Scoggan 1996; Taylor 1996), a pesar de su cobertura en decenas de libros y artículos populares, y el hecho de que la interpretación del plesiosauro a menudo se presenta de una manera  más favorable que la evidencia científica.
más favorable que la evidencia científica.
Recientemente, los creacionistas han escrito dos resúmenes más exactos del caso, pero aún incompletos. Niermann (1994) señaló que los estudios de 1978 se referían a un tiburón, el tiburón peregrino, y que tendía a descomponerse en formas parecidas a un plesiosauro.  Lamentablemente, oculta estos comentarios en notas a pie de página, mientras que el cuerpo del texto se alienta la interpretación del plesiosauro. Todd Wood (1997) reconoció que la evidencia respalda fuertemente la conclusión del tiburón peregrino, pero enumera varias supuestas incoherencias con la identificación con tiburones – ninguna de las cuales analizaremos exhaustivamente.
Lamentablemente, oculta estos comentarios en notas a pie de página, mientras que el cuerpo del texto se alienta la interpretación del plesiosauro. Todd Wood (1997) reconoció que la evidencia respalda fuertemente la conclusión del tiburón peregrino, pero enumera varias supuestas incoherencias con la identificación con tiburones – ninguna de las cuales analizaremos exhaustivamente.
 Como era de esperar, la historia del monstruo de Nueva Zelanda también ha derivado en la Internet. Los creacionistas Kent Hovind (1996), Walter Brown (1996), Bernard Northrup (1997), Paul Smithson (1996), y Don Patton (1995) alientan la interpretación del plesiosauro. Brown llama a la criatura un plesiosauro, que él incorrectamente llama un «dinosaurio» marino. También señala que el cadáver tenía
Como era de esperar, la historia del monstruo de Nueva Zelanda también ha derivado en la Internet. Los creacionistas Kent Hovind (1996), Walter Brown (1996), Bernard Northrup (1997), Paul Smithson (1996), y Don Patton (1995) alientan la interpretación del plesiosauro. Brown llama a la criatura un plesiosauro, que él incorrectamente llama un «dinosaurio» marino. También señala que el cadáver tenía  vértebras, afirmando que se trata del «algo que no está presente en muchos peces, incluidos los tiburones». (Por supuesto, los peces, incluidos los tiburones, tienen vértebras). En cambio, el sitio web de Strange Magazine ofrece bastantes buenos resúmenes del canal del Zuiyo-Maru y otros varios canales varados, al igual que Roesch (1997a).
vértebras, afirmando que se trata del «algo que no está presente en muchos peces, incluidos los tiburones». (Por supuesto, los peces, incluidos los tiburones, tienen vértebras). En cambio, el sitio web de Strange Magazine ofrece bastantes buenos resúmenes del canal del Zuiyo-Maru y otros varios canales varados, al igual que Roesch (1997a).
Después de todo, incluso entre un grupo de pescadores se hubieran dado cuenta de que un «monstruo marino» prehistórico sería de mayor valor más económico y científico que una carga de peces.
Continuará…
[1] Kuban J. Glen, Sea Monster or Shark? An Analysis of a Supposed Plesiosaur Carcass Netted in 1977, Reports of the National Center for Science Education, Vol. 17, No. 3, 1997, Pags. 16«“28, http://paleo.cc/paluxy/plesios.htm
esta pagina es chevverrere se la voy a mandar amis amigos
una buena investigación que definitivamente debería tener mayor difusión